

| 產品編號 | bs-12412R |
| 英文名稱 | phospho-SP1 (Thr453) Rabbit pAb |
| 中文名稱 | 磷酸化轉錄生長因子SP1抗體 |
| 別 名 | SP1(phospho T453); p-SP1(phospho T453); SP1(phospho-Thr453); SP1(phospho-T453); p-SP1(phospho T453); p-TSFP1(phospho T453); Sp1 transcription factor isoform a; TSFP1; TSFP 1; Specificity protein 1; Transcription factor Sp1; SP1_HUMAN. |
 |
Specific References (1) | bs-12412R has been referenced in 1 publications.
[IF=2.523] Zhang C et al. Caveolin-1 promotes Rfng expression via Erk-Jnk-p38 signaling pathway in mouse hepatocarcinoma cells. J Physiol Biochem. 2019 Sep 16. WB ; Mouse.
|
| 產品類型 | 磷酸化抗體 |
| 研究領域 | 腫瘤 細胞生物 發育生物學 神經生物學 信號轉導 干細胞 轉錄調節因子 鋅指蛋白 表觀遺傳學 |
| 抗體來源 | Rabbit |
| 克隆類型 | Polyclonal |
| 交叉反應 | Human (predicted: Mouse,Rat) |
| 產品應用 | IHC-P=1:100-500,IHC-F=1:100-500,IF=1:100-500
not yet tested in other applications. optimal dilutions/concentrations should be determined by the end user. |
| 理論分子量 | 81 kDa |
| 檢測分子量 | |
| 細胞定位 | 細胞核 細胞漿 |
| 性 狀 | Liquid |
| 濃 度 | 1mg/ml |
| 免 疫 原 | KLH conjugated synthesised phosphopeptide derived from human SP1 around the phosphorylation site of Thr453: IR(p-T)PT |
| 亞 型 | IgG |
| 純化方法 | affinity purified by Protein A |
| 緩 沖 液 | 0.01M TBS (pH7.4) with 1% BSA, 0.02% Proclin300 and 50% Glycerol. |
| 保存條件 | Shipped at 4℃. Store at -20℃ for one year. Avoid repeated freeze/thaw cycles. |
| 注意事項 | This product as supplied is intended for research use only, not for use in human, therapeutic or diagnostic applications. |
| PubMed | PubMed |
| 產品介紹 |
Sp1 is a sequence-specific transcription factor that recognizes GGGGCGGGGC and closely related sequences, which are often referred to as GC boxes. Sp1 was initially identified as a HeLa cell-derived factor that selectively activates in vitro transcription from the SV40 promoter and binds to the multiple GC boxes in the 21-bp repeated elements in SV40. The sequence specificity of DNA binding is conferred by Zn (II) fingers, whereas a different region of Sp1 appears to regulate the affinity of DNA binding. Sp1 belongs to a subgroup of transcription factors that are phosphorylated upon binding to promoter sequences. Evidence suggests that the early growth response gene, Erg-1 (also known as Zif268 or NGF1-A) (7), may downregulate certain mammalian gene promoters by competing with Sp1 for binding to an overlapping binding motif. The gene encoding human Sp1 maps to chromosome 12q13.1. Function: Transcription factor that can activate or repress transcription in response to physiological and pathological stimuli. Binds with high affinity to GC-rich motifs and regulates the expression of a large number of genes involved in a variety of processes such as cell growth, apoptosis, differentiation and immune responses. Highly regulated by post-translational modifications (phosphorylations, sumoylation, proteolytic cleavage, glycosylation and acetylation). Binds also the PDGFR-alpha G-box promoter. May have a role in modulating the cellular response to DNA damage. Implicated in chromatin remodeling. Plays a role in the recruitment of SMARCA4/BRG1 on the c-FOS promoter. Plays an essential role in the regulation of FE65 gene expression. In complex with ATF7IP, maintains telomerase activity in cancer cells by inducing TERT and TERC gene expression. Subunit: Interacts with ATF7IP, ATF7IP2, BAHD1, POGZ, HCFC1, AATF and PHC2. Interacts with varicella-zoster virus IE62 protein. Interacts with HIV-1 Vpr; the interaction is inhibited by SP1 O-glycosylation. Interacts with SV40 VP2/3 proteins. Interacts with SV40 major capsid protein VP1; this interaction leads to a cooperativity between the 2 proteins in DNA binding. Interacts with HLTF; the interaction may be required for basal transcriptional activity of HLTF. Interacts (deacetylated form) with EP300; the interaction enhances gene expression. Interacts with HDAC1 and JUN. Interacts with ELF1; the interaction is inhibited by glycosylation of SP1. Interaction with NFYA; the interaction is inhibited by glycosylation of SP1. Interacts with SMARCA4/BRG1. Interacts with ATF7IP and TBP. Interacts with MEIS2 isoform 4 and PBX1 isoform PBX1a. Subcellular Location: Nucleus. Cytoplasm. Nuclear location is governed by glycosylated/phosphorylated states. Insulin promotes nuclear location, while glucagon favors cytoplasmic location. Tissue Specificity: Up-regulated in adenocarcinomas of the stomach (at protein level). Post-translational modifications: Phosphorylated on multiple serine and threonine residues. Phosphorylation is coupled to ubiquitination, sumoylation and proteolytic processing. Phosphorylation on Ser-59 enhances proteolytic cleavage. Phosphorylation on Ser-7 enhances ubiquitination and protein degradation. Hyperphosphorylation on Ser-101 in response to DNA damage has no effect on transcriptional activity. MAPK1/MAPK3-mediated phosphorylation on Thr-453 and Thr-739 enhances VEGF transcription but, represses FGF2-triggered PDGFR-alpha transcription. Also implicated in the repression of RECK by ERBB2. Hyperphosphorylated on Thr-278 and Thr-739 during mitosis by MAPK8 shielding SP1 from degradation by the ubiquitin-dependent pathway. Phosphorylated in the zinc-finger domain by calmodulin-activated PKCzeta. Phosphorylation on Ser-641 by PKCzeta is critical for TSA-activated LHR gene expression through release of its repressor, p107. Phosphorylation on Thr-668, Ser-670 and Thr-681 is stimulated by angiotensin II via the AT1 receptor inducing increased binding to the PDGF-D promoter. This phosphorylation is increased in injured artey wall. Ser-59 and Thr-681 can both be dephosphorylated by PP2A during cell-cycle interphase. Dephosphorylation on Ser-59 leads to increased chromatin association during interphase and increases the transcriptional activity. On insulin stimulation, sequentially glycosylated and phosphorylated on several C-terminal serine and threonine residues. Acetylated. Acetylation/deacetylation events affect transcriptional activity. Deacetylation leads to an increase in the expression the 12(s)-lipooxygenase gene though recruitment of p300 to the promoter. Ubiquitinated. Ubiquitination occurs on the C-terminal proteolytically-cleaved peptide and is triggered by phosphorylation. Sumoylated by SUMO1. Sumoylation modulates proteolytic cleavage of the N-terminal repressor domain. Sumoylation levels are attenuated during tumorigenesis. Phosphorylation mediates SP1 desumoylation. Proteolytic cleavage in the N-terminal repressor domain is prevented by sumoylation. The C-terminal cleaved product is susceptible to degradation. O-glycosylated; contains at least 8 N-acetylglucosamine side chains. Levels are controlled by insulin and the SP1 phosphorylation states. Insulin-mediated O-glycosylation locates SP1 to the nucleus, where it is sequentially deglycosylated and phosphorylated. O-glycosylation affects transcriptional activity through disrupting the interaction with a number of transcription factors including ELF1 and NFYA. Also inhibits interaction with the HIV1 promoter. Inhibited by peroxisomome proliferator receptor gamma (PPARgamma). Similarity: Belongs to the Sp1 C2H2-type zinc-finger protein family. Contains 3 C2H2-type zinc fingers. SWISS: P08047 Gene ID: 6667 Database links: Entrez Gene: 6667 Human Entrez Gene: 20683 Mouse Omim: 189906 Human SwissProt: P08047 Human SwissProt: O89090 Mouse |
| 產品圖片 |
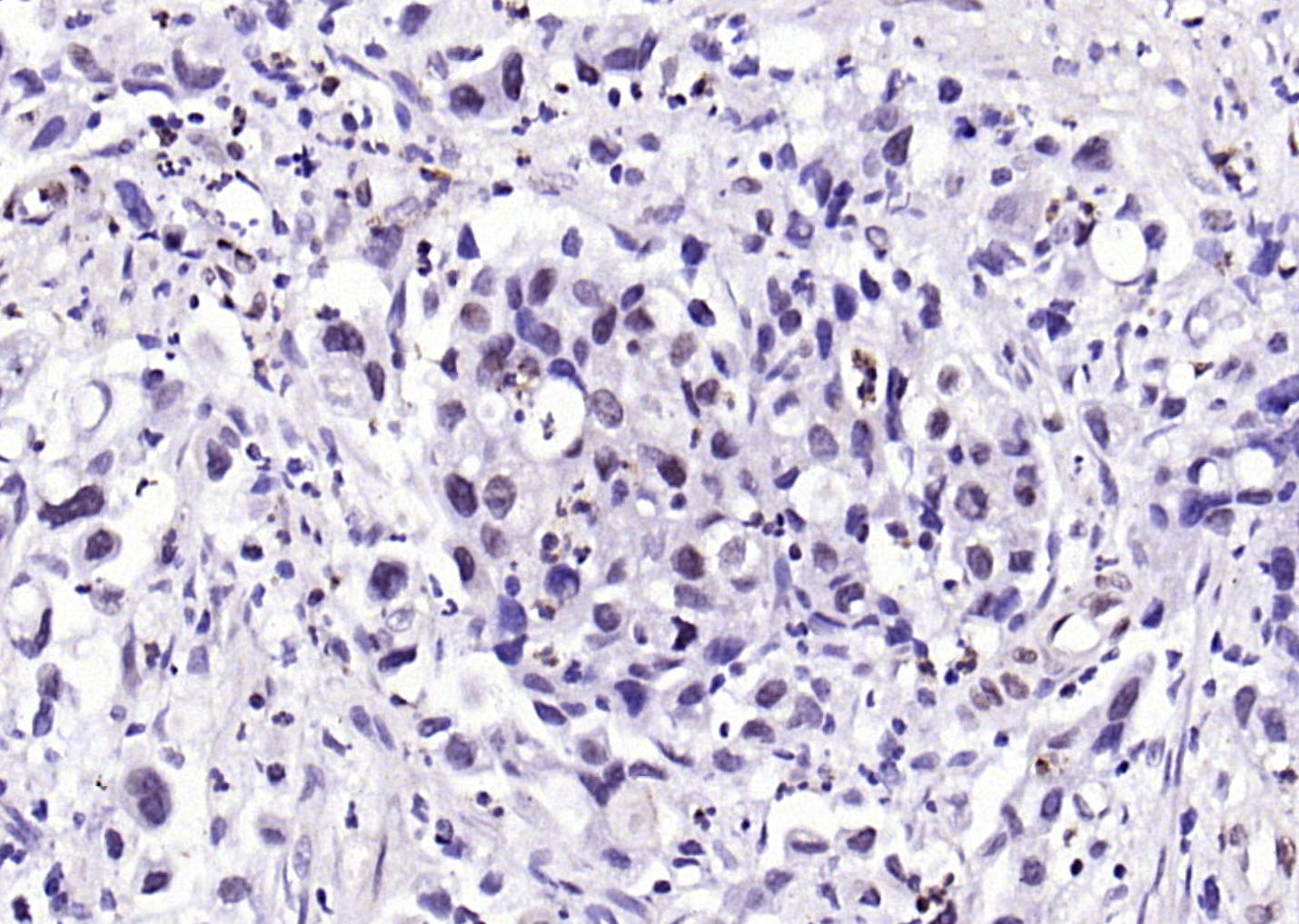
Paraformaldehyde-fixed, paraffin embedded (human gastric carcinoma); Antigen retrieval by boiling in sodium citrate buffer (pH6.0) for 15min; Block endogenous peroxidase by 3% hydrogen peroxide for 20 minutes; Blocking buffer (normal goat serum) at 37°C for 30min; Antibody incubation with (phospho-SP1 (Thr453)) Polyclonal Antibody, Unconjugated (bs-12412R) at 1:200 overnight at 4°C, followed by operating according to SP Kit(Rabbit) (sp-0023) instructionsand DAB staining.
|
| 1、抗體溶解方法 | |
| 2、抗體修復方式 | |
| 3、常用試劑的配制 | |
| 4、免疫組化操作步驟 | |
| 5、免疫組化問題解答 | |
| 6、Western Blotting 操作步驟 | |
| 7、Western Blotting 問題解答 | |
| 8、關于肽鏈的設計 | |
| 9、多肽的溶解與保存 | |
| 10、酶標抗體效價測定程序 | |





