
| 產品編號 | bs-3484R |
| 英文名稱 | Rabbit Anti-Smad3 antibody |
| 中文名稱 | 細胞信號轉導分子SMAD3抗體 |
| 別 名 | hMAD 3; hSMAD3; HSPC193; JV15 2; JV152; MAD (mothers against decapentaplegic Drosophila) homolog 3; MAD3; MADH 3; MADH3; Mothers against decapentaplegic homolog 3; Mothers against DPP homolog 3; SMA and MAD related protein 3; SMAD 3; SMAD; SMAD-3; SMAD3_HUMAN. |
 |
Specific References (33) | bs-3484R has been referenced in 33 publications.
|
| 研究領域 | 腫瘤 細胞生物 免疫學 信號轉導 干細胞 細胞凋亡 生長因子和激素 轉錄調節因子 |
| 抗體來源 | Rabbit |
| 克隆類型 | Polyclonal |
| 交叉反應 | Human,Mouse,Rat,Pig (predicted: Cow,Chicken) |
| 產品應用 | WB=1:500-2000,IHC-P=1:100-500,IHC-F=1:100-500,Flow-Cyt=1ug/Test,ICC/IF=1:100,IF=1:100-500,ELISA=1:5000-10000
not yet tested in other applications. optimal dilutions/concentrations should be determined by the end user. |
| 理論分子量 | 47kDa |
| 細胞定位 | 細胞核 細胞漿 |
| 性 狀 | Liquid |
| 濃 度 | 1mg/ml |
| 免 疫 原 | KLH conjugated synthetic peptide derived from human Smad3: 31-80/425 |
| 亞 型 | IgG |
| 純化方法 | affinity purified by Protein A |
| 緩 沖 液 | 0.01M TBS (pH7.4) with 1% BSA, 0.02% Proclin300 and 50% Glycerol. |
| 保存條件 | Shipped at 4℃. Store at -20℃ for one year. Avoid repeated freeze/thaw cycles. |
| 注意事項 | This product as supplied is intended for research use only, not for use in human, therapeutic or diagnostic applications. |
| PubMed | PubMed |
| 產品介紹 |
Smad3 is a 50 kDa member of a family of proteins that act as key mediators of TGF beta superfamily signaling in cell proliferation, differentiation and development. The Smad family is divided into three subclasses: receptor regulated Smads, activin/TGF beta receptor regulated (Smad2 and 3) or BMP receptor regulated (Smad 1, 5, and 8); the common partner, (Smad4) that functions via its interaction to the various Smads; and the inhibitory Smads, (Smad6 and 7). Activated Smad3 oligomerizes with Smad4 upon TGF beta stimulation and translocates as a complex into the nucleus, allowing its binding to DNA and transcription factors. Phosphorylation of the two TGF beta dependent serines 423 and 425 in the C terminus of Smad3 is critical for Smad3 transcriptional activity and TGF beta signaling. Function: Receptor-regulated SMAD (R-SMAD) that is an intracellular signal transducer and transcriptional modulator activated by TGF-beta (transforming growth factor) and activin type 1 receptor kinases. Binds the TRE element in the promoter region of many genes that are regulated by TGF-beta and, on formation of the SMAD3/SMAD4 complex, activates transcription. Also can form a SMAD3/SMAD4/JUN/FOS complex at the AP-1/SMAD site to regulate TGF-beta-mediated transcription. Has an inhibitory effect on wound healing probably by modulating both growth and migration of primary keratinocytes and by altering the TGF-mediated chemotaxis of monocytes. This effect on wound healing appears to be hormone-sensitive. Regulator of chondrogenesis and osteogenesis and inhibits early healing of bone fractures (By similarity). Positively regulates PDPK1 kinase activity by stimulating its dissociation from the 14-3-3 protein YWHAQ which acts as a negative regulator. Subunit: Monomer; in the absence of TGF-beta. Homooligomer; in the presence of TGF-beta. Heterotrimer; forms a heterotrimer in the presence of TGF-beta consisting of two molecules of C-terminally phosphorylated SMAD2 or SMAD3 and one of SMAD4 to form the transcriptionally active SMAD2/SMAD3-SMAD4 complex. Interacts with TGFBR1. Part of a complex consisting of AIP1, ACVR2A, ACVR1B and SMAD3. Interacts with AIP1, TGFB1I1, TTRAP, FOXL2, PML, PRDM16, HGS and WWP1. Interacts (via MH2 domain) with CITED2 (via C-terminus) (By similarity). Interacts with NEDD4L; the interaction requires TGF-beta stimulation (By similarity). Interacts (via the MH2 domain) with ZFYVE9. Interacts with HDAC1, VDR, TGIF and TGIF2, RUNX3, CREBBP, SKOR1, SKOR2, SNON, ATF2, SMURF2 and TGFB1I1. Interacts with DACH1; the interaction inhibits the TGF-beta signaling. Forms a complex with SMAD2 and TRIM33 upon addition of TGF-beta. Found in a complex with SMAD3, RAN and XPO4. Interacts in the complex directly with XPO4. Interacts (via the MH2 domain) with LEMD3; the interaction represses SMAD3 transcriptional activity through preventing the formation of the heteromeric complex with SMAD4 and translocation to the nucleus. Interacts with RBPMS. Interacts (via MH2 domain) with MECOM. Interacts with WWTR1 (via its coiled-coil domain). Interacts (via the linker region) with EP300 (C-terminal); the interaction promotes SMAD3 acetylation and is enhanced by TGF-beta phosphorylation in the C-terminal of SMAD3. This interaction can be blocked by competitive binding of adenovirus oncoprotein E1A to the same C-terminal site on EP300, which then results in partially inhibited SMAD3/SMAD4 transcriptional activity. Interacts with SKI; the interaction represses SMAD3 transcriptional activity. Component of the multimeric complex SMAD3/SMAD4/JUN/FOS which forms at the AP1 promoter site; required for syngernistic transcriptional activity in response to TGF-beta. Interacts (via an N-terminal domain) with JUN (via its basic DNA binding and leucine zipper domains); this interaction is essential for DNA binding and cooperative transcriptional activity in response to TGF-beta. Interacts with PPM1A; the interaction dephosphorylates SMAD3 in the C-terminal SXS motif leading to disruption of the SMAD2/3-SMAD4 complex, nuclear export and termination of TGF-beta signaling. Interacts (dephosphorylated form via the MH1 and MH2 domains) with RANBP3 (via its C-terminal R domain); the interaction results in the export of dephosphorylated SMAD3 out of the nucleus and termination of the TGF-beta signaling. Interacts with MEN1. Interacts with IL1F7. Interaction with CSNK1G2. Interacts with PDPK1 (via PH domain). Subcellular Location: Cytoplasm. Nucleus. Note=Cytoplasmic and nuclear in the absence of TGF-beta. On TGF-beta stimulation, migrates to the nucleus when complexed with SMAD4. Through the action of the phosphatase PPM1A, released from the SMAD2/SMAD4 complex, and exported out of the nucleus by interaction with RANBP1. Co-localizes with LEMD3 at the nucleus inner membrane. MAPK-mediated phosphorylation appears to have no effect on nuclear import. PDPK1 prevents its nuclear translocation in response to TGF-beta. Post-translational modifications: Phosphorylated on serine and threonine residues. Enhanced phosphorylation in the linker region on Thr-179, Ser-204 and Ser-208 on EGF AND TGF-beta treatment. Ser-208 is the main site of MAPK-mediated phosphorylation. CDK-mediated phosphorylation occurs in a cell-cycle dependent manner and inhibits both the transcriptional activity and antiproliferative functions of SMAD3. This phosphorylation is inhibited by flavopiridol. Maximum phosphorylation at the G(1)/S junction. Also phosphorylated on serine residues in the C-terminal SXS motif by TGFBR1 and ACVR1. TGFBR1-mediated phosphorylation at these C-terminal sites is required for interaction with SMAD4, nuclear location and transactivational activity, and appears to be a prerequisite for the TGF-beta mediated phosphorylation in the linker region. Dephosphorylated in the C-terminal SXS motif by PPM1A. This dephosphorylation disrupts the interaction with SMAD4, promotes nuclear export and terminates TGF-beta-mediated signaling. Phosphorylation at Ser-418 by CSNK1G2/CK1 promotes ligand-dependent ubiquitination and subsequent proteasome degradation, thus inhibiting SMAD3-mediated TGF-beta responses. Phosphorylated by PDPK1. Acetylation in the nucleus by EP300 in the MH2 domain regulates positively its transcriptional activity and is enhanced by TGF-beta. Ubiquitinated. DISEASE: Defects in SMAD3 may be a cause of colorectal cancer (CRC) [MIM:114500]. Defects in SMAD3 are the cause of Loeys-Dietz syndrome type 1C (LDS1C) [MIM:613795]. LDS1C is an aortic aneurysm syndrome with widespread systemic involvement. The disorder is characterized by the triad of arterial tortuosity and aneurysms, hypertelorism, and bifid uvula or cleft palate. Patients with LDS1C also manifest early-onset osteoarthritis. They lack craniosynostosis and mental retardation. Similarity: Belongs to the dwarfin/SMAD family. Contains 1 MH1 (MAD homology 1) domain. Contains 1 MH2 (MAD homology 2) domain. SWISS: Q15796 Gene ID: 4088 Database links: Entrez Gene: 4088 Human Entrez Gene: 17127 Mouse Omim: 603109 Human SwissProt: P84022 Human SwissProt: Q8BUN5 Mouse Unigene: 727986 Human Unigene: 7320 Mouse Unigene: 10636 Rat |
| 產品圖片 |
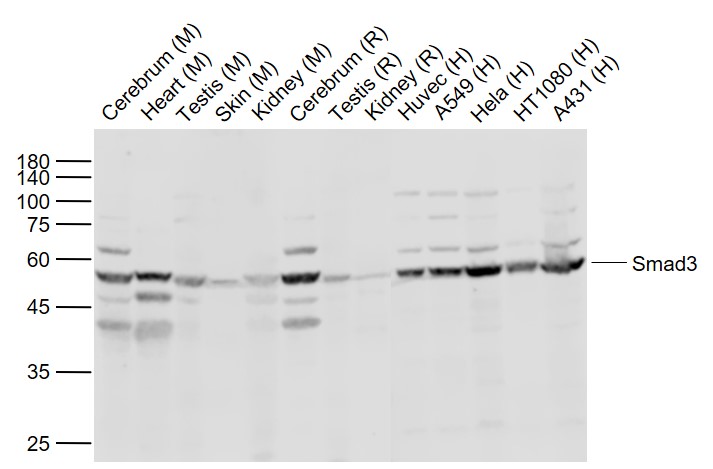
Sample:
Lane 1: Cerebrum (Mouse) Lysate at 40 ug
Lane 2: Heart (Mouse) Lysate at 40 ug
Lane 3: Testis (Mouse) Lysate at 40 ug
Lane 4: Skin (Mouse) Lysate at 40 ug
Lane 5: Kidney (Mouse) Lysate at 40 ug
Lane 6: Cerebrum (Rat) Lysate at 40 ug
Lane 7: Testis (Rat) Lysate at 40 ug
Lane 8: Kidney (Rat) Lysate at 40 ug
Lane 9: Huvec (Human) Cell Lysate at 30 ug
Lane 10: A549 (Human) Cell Lysate at 30 ug
Lane 11: Hela (Human) Cell Lysate at 30 ug
Lane 12: HT1080 (Human) Cell Lysate at 30 ug
Lane 13: A431 (Human) Cell Lysate at 30 ug
Primary: Anti-Smad3 (bs-3484R) at 1/1000 dilution
Secondary: IRDye800CW Goat Anti-Rabbit IgG at 1/20000 dilution
Predicted band size: 52 kD
Observed band size: 54 kD
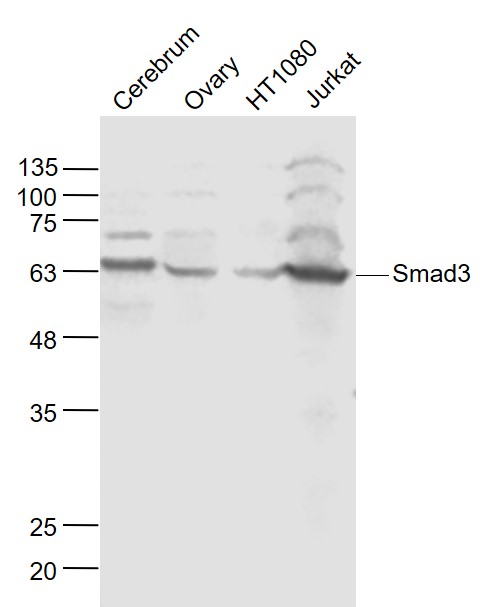
Sample:
Cerebrum (Mouse) Lysate at 40 ug
Ovary (Mouse) Lysate at 40 ug
HT1080 (Human) Cell Lysate at 30 ug
Jurkat (Human) Cell Lysate at 30 ug
Primary: Anti- Smad3 (bs-3484R) at 1/1000 dilution
Secondary: IRDye800CW Goat Anti-Rabbit IgG at 1/20000 dilution
Predicted band size: 52 kD
Observed band size: 60 kD
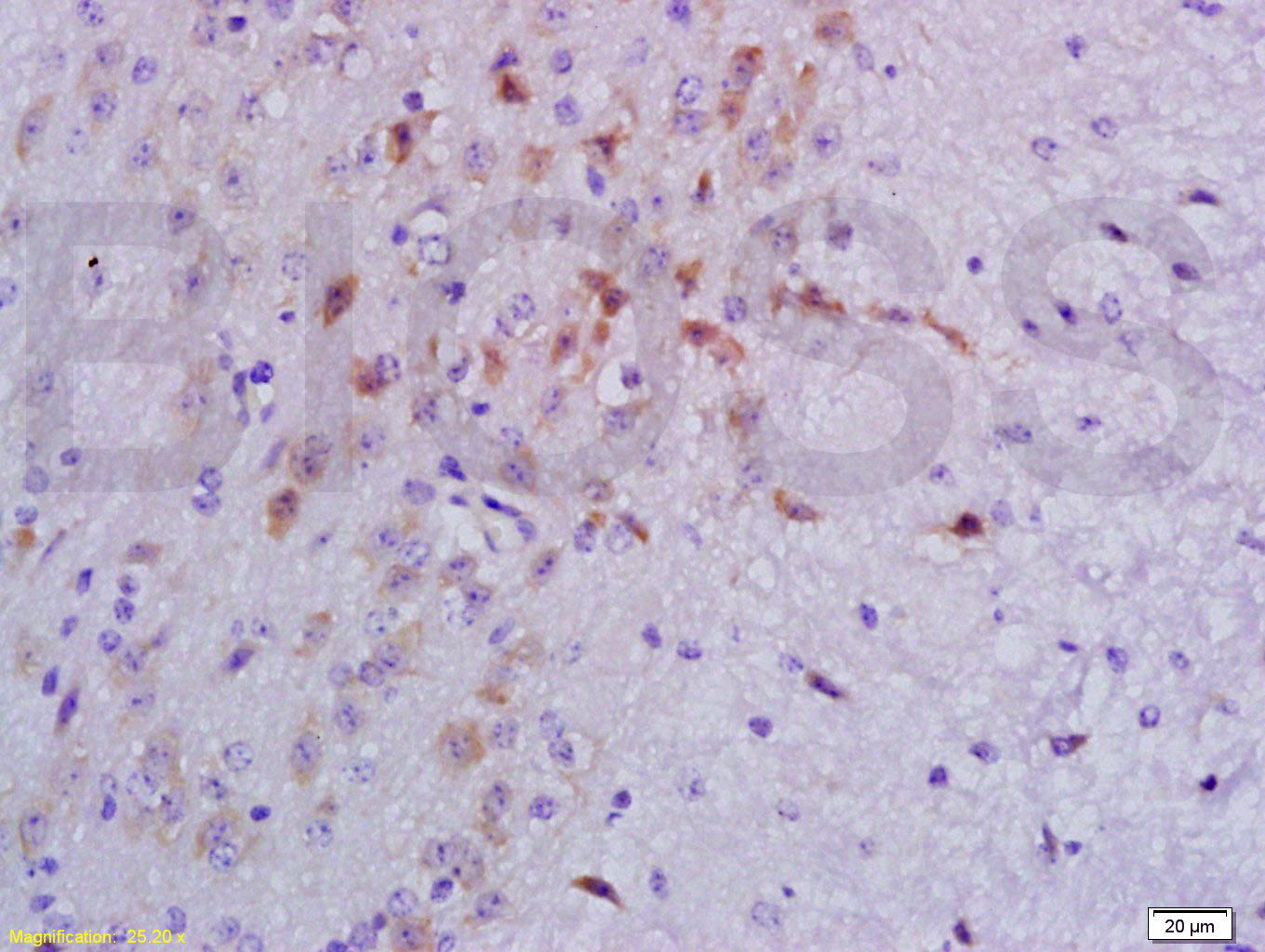
Tissue/cell: mouse lymphoma tissue; 4% Paraformaldehyde-fixed and paraffin-embedded;
Antigen retrieval: citrate buffer ( 0.01M, pH 6.0 ), Boiling bathing for 15min; Block endogenous peroxidase by 3% Hydrogen peroxide for 30min; Blocking buffer (normal goat serum,C-0005) at 37℃ for 20 min;
Incubation: Anti-Smad3 Polyclonal Antibody, Unconjugated(bs-3484R) 1:200, overnight at 4°C, followed by conjugation to the secondary antibody(SP-0023) and DAB(C-0010) staining
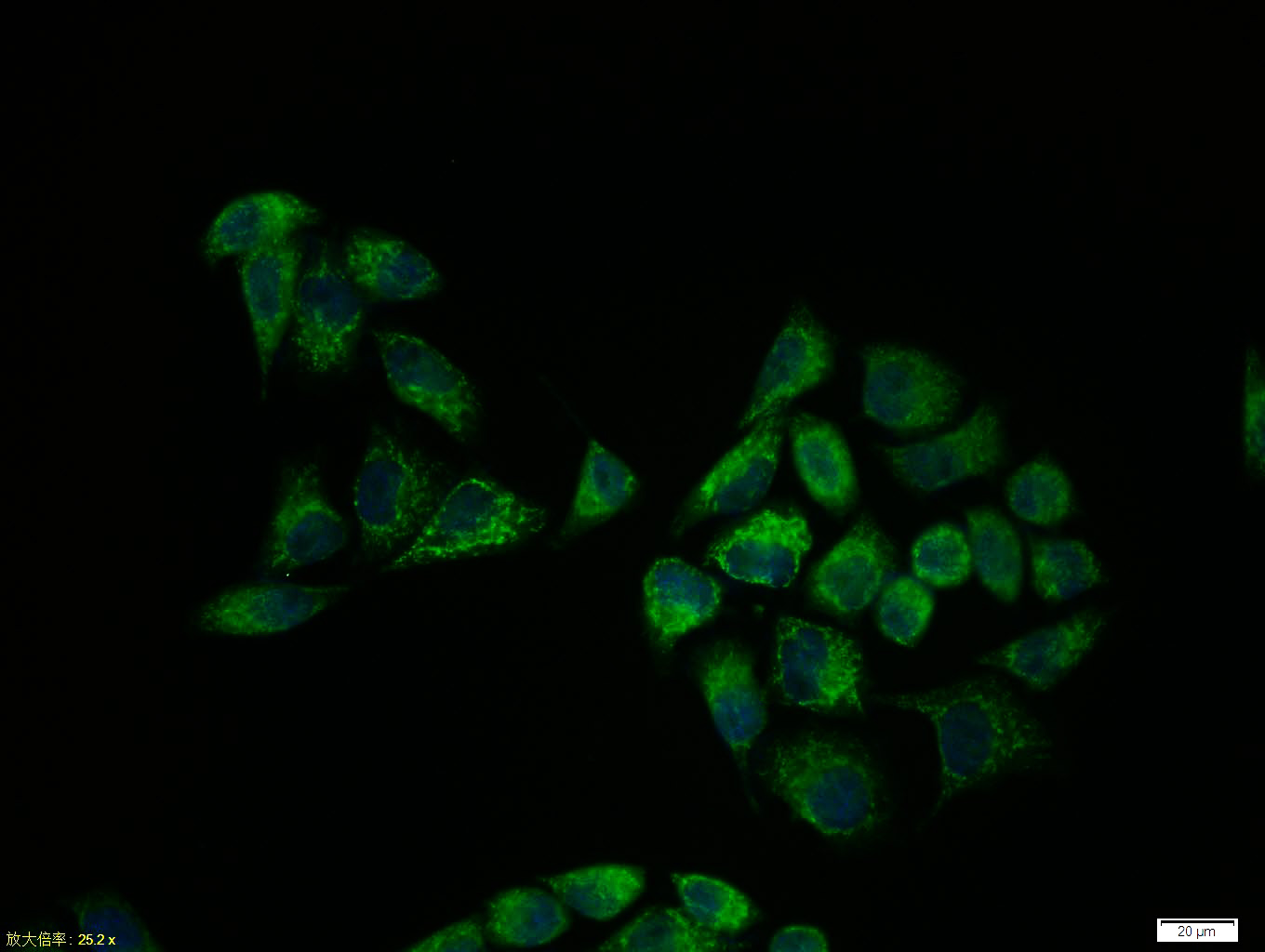
Hela cell; 4% Paraformaldehyde-fixed; Triton X-100 at room temperature for 20 min; Blocking buffer (normal goat serum, C-0005) at 37°C for 20 min; Antibody incubation with (Smad3) polyclonal Antibody, Unconjugated (bs-3484R) 1:100, 90 minutes at 37°C; followed by a conjugated Goat Anti-Rabbit IgG antibody at 37°C for 90 minutes, DAPI (blue, C02-04002) was used to stain the cell nuclei.
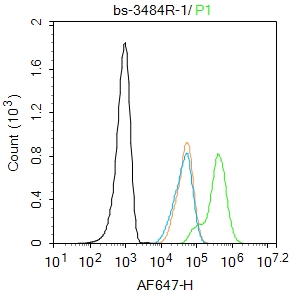
Blank control: Hela.
Primary Antibody (green line): Rabbit Anti-Smad3 antibody (bs-3484R)
Dilution: 1μg /10^6 cells;
Isotype Control Antibody (orange line): Rabbit IgG .
Secondary Antibody : Goat anti-rabbit IgG-AF647
Dilution: 1μg /test.
Protocol
The cells were fixed with 4% PFA (10min at room temperature)and then permeabilized with 90% ice-cold methanol for 20 min at -20℃. The cells were then incubated in 5%BSA to block non-specific protein-protein interactions for 30 min at room temperature .Cells stained with Primary Antibody for 30 min at room temperature. The secondary antibody used for 40 min at room temperature. Acquisition of 20,000 events was performed.
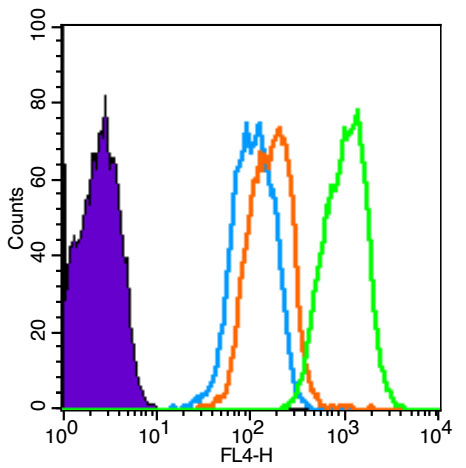
Blank control (Black line): HUVEC (Black).
Primary Antibody (green line): Rabbit Anti-Smad3 antibody (bs-3484R)
Dilution: 1μg /10^6 cells;
Isotype Control Antibody (orange line): Rabbit IgG .
Secondary Antibody (white blue line): Goat anti-rabbit IgG-AF647
Dilution: 1μg /test.
Protocol
The cells were fixed with 4% PFA (10min at room temperature)and then permeabilized with 90% ice-cold methanol for 20 min at room temperature. The cells were then incubated in 5%BSA to block non-specific protein-protein interactions for 30 min at room temperature .Cells stained with Primary Antibody for 30 min at room temperature. The secondary antibody used for 40 min at room temperature. Acquisition of 20,000 events was performed.
|
| 1、抗體溶解方法 | |
| 2、抗體修復方式 | |
| 3、常用試劑的配制 | |
| 4、免疫組化操作步驟 | |
| 5、免疫組化問題解答 | |
| 6、Western Blotting 操作步驟 | |
| 7、Western Blotting 問題解答 | |
| 8、關于肽鏈的設計 | |
| 9、多肽的溶解與保存 | |
| 10、酶標抗體效價測定程序 | |






